Macrophage polarization is a central focus in immunological research, and accurate detection of its functional subtypes—such as pro-inflammatory M1 and anti-inflammatory M2—forms the basis for deciphering immune regulation mechanisms and conducting drug screening. In our previous article "Macrophage Function and Detection Techniques | Macrophage Polarization: Mechanisms, Protocols, and Key Experimental Considerations", we outlined polarization classification, experimental protocols, and essential quality-control points, providing researchers with a standardized experimental framework.
In actual research practice, however, a frustrating phenomenon is frequently encountered: even when using the same cell line, identical concentrations of polarizing agents (e.g., LPS+IFN-γ for M1, IL 4 for M2a), and detecting well-established polarization markers, results often vary between different laboratories or even between different experimental batches within the same lab. Some experiments clearly capture high marker expression, while others show "false negatives," leaving researchers perplexed. What causes such discrepancies? Beyond common factors like cell status and reagent purity, polarization time may be an overlooked variable.
A 2022 study "Effect of stimulation time on the expression of human macrophage polarization markers" systematically addressed this issue using human monocyte-derived macrophages (MDMs) as a model. Through time-course analysis, the study demonstrated that stimulation time significantly affects the expression intensity and peak profiles of polarization markers. This finding offers a fresh perspective for interpreting experimental variability and provides a new direction to consider when experiments reach an impasse.
I. Literature Focus: A Targeted Study Exploring the Potential Impact of Time
To clarify the relationship between time and polarization detection, the research team employed human monocyte- derived macrophages (MDMs)—a model closer to human physiology that avoids interference from differences between cell lines like THP-1 and primary cells—and designed a systematic controlled experiment specifically examining the temporal effect on polarization markers:
Experimental Design
The core value of this study lies not in simply confirming "whether polarization occurs," but in systematically presenting the dynamic trajectories of different markers across subtypes and detection dimensions over time, providing direct evidence for understanding why timing matters.
II. Key Findings: Temporal Dynamics Reshape Polarization Marker Expression
Results revealed that both different markers within the same polarization subtype and similar markers across different subtypes exhibit distinct "temporal rhythms"—likely a major contributor to experimental variability.
1. M1 Macrophages (Pro-inflammatory Subtype): Marker Expression Follows Distinct Timelines
As the classic pro-inflammatory subtype, M1 markers are not synchronously activated but display clear time-layered characteristics:
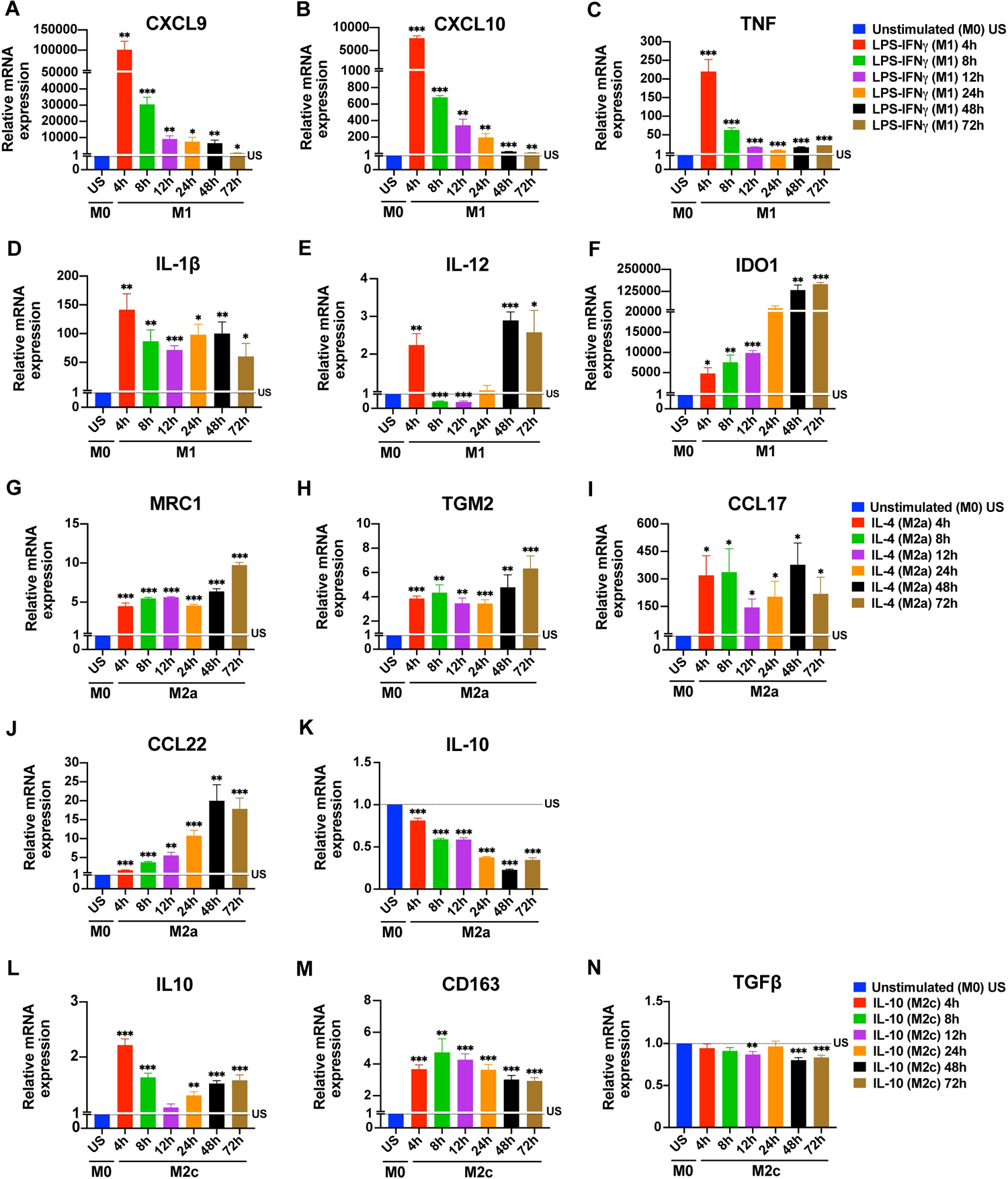
Figure 1. Time-dependent changes in the expression of macrophage polarization markers at mRNA
2. M2a Macrophages (Repair Subtype): "Slow-Burn" Expression Profile
M2a macrophages, involved in tissue repair and wound healing, generally exhibit a slower, more gradual marker upregulation compared to M1:
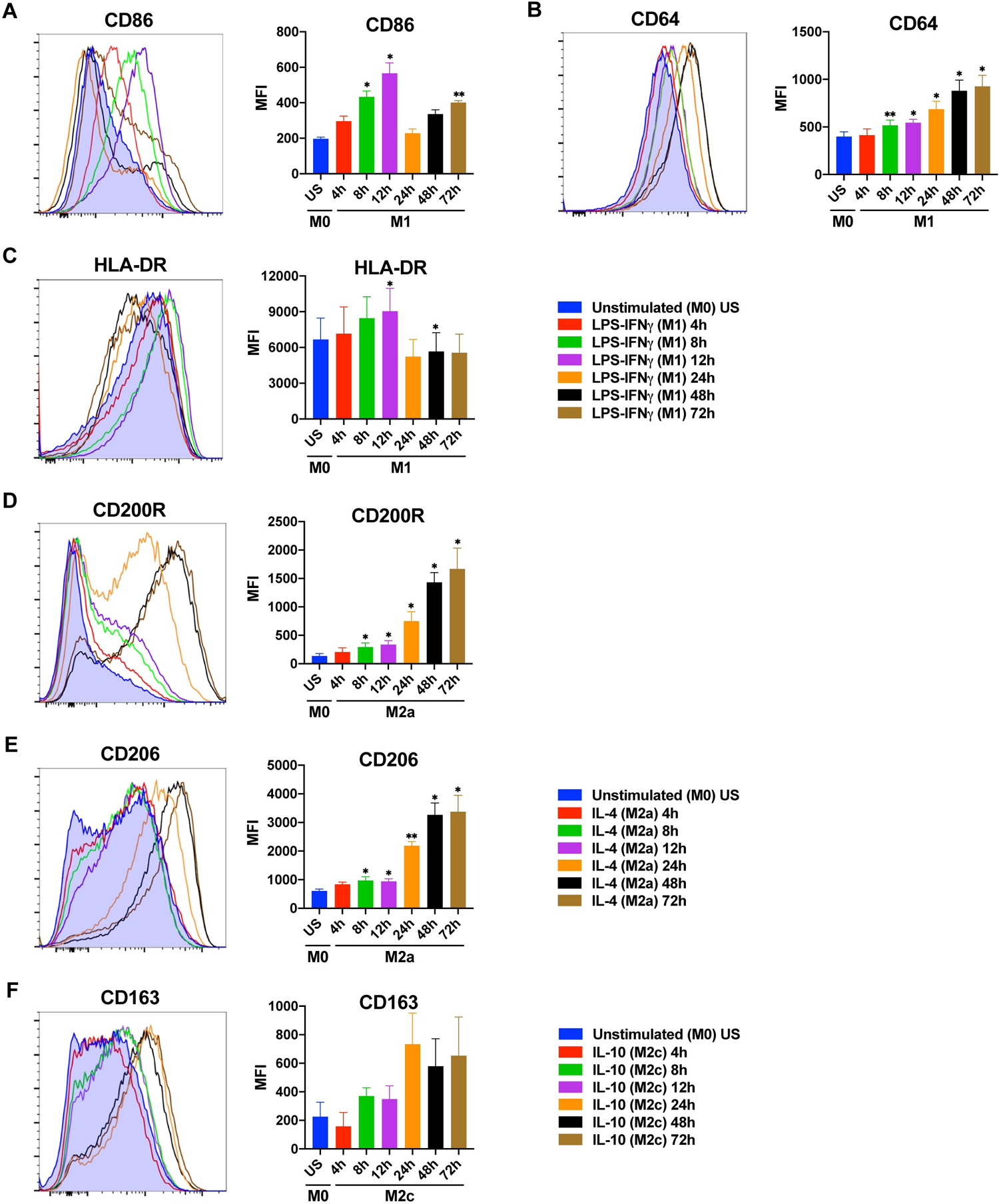
Figure 2. Time-dependent changes in the expression of macrophage polarization markers at protein
3. M2c Macrophages (Immunoregulatory Subtype): Transcription-Secretion "Time Lag"
Induced by IL-10, M2c macrophages are associated with anti-inflammation and immune tolerance, and their temporal patterns are particularly distinctive:
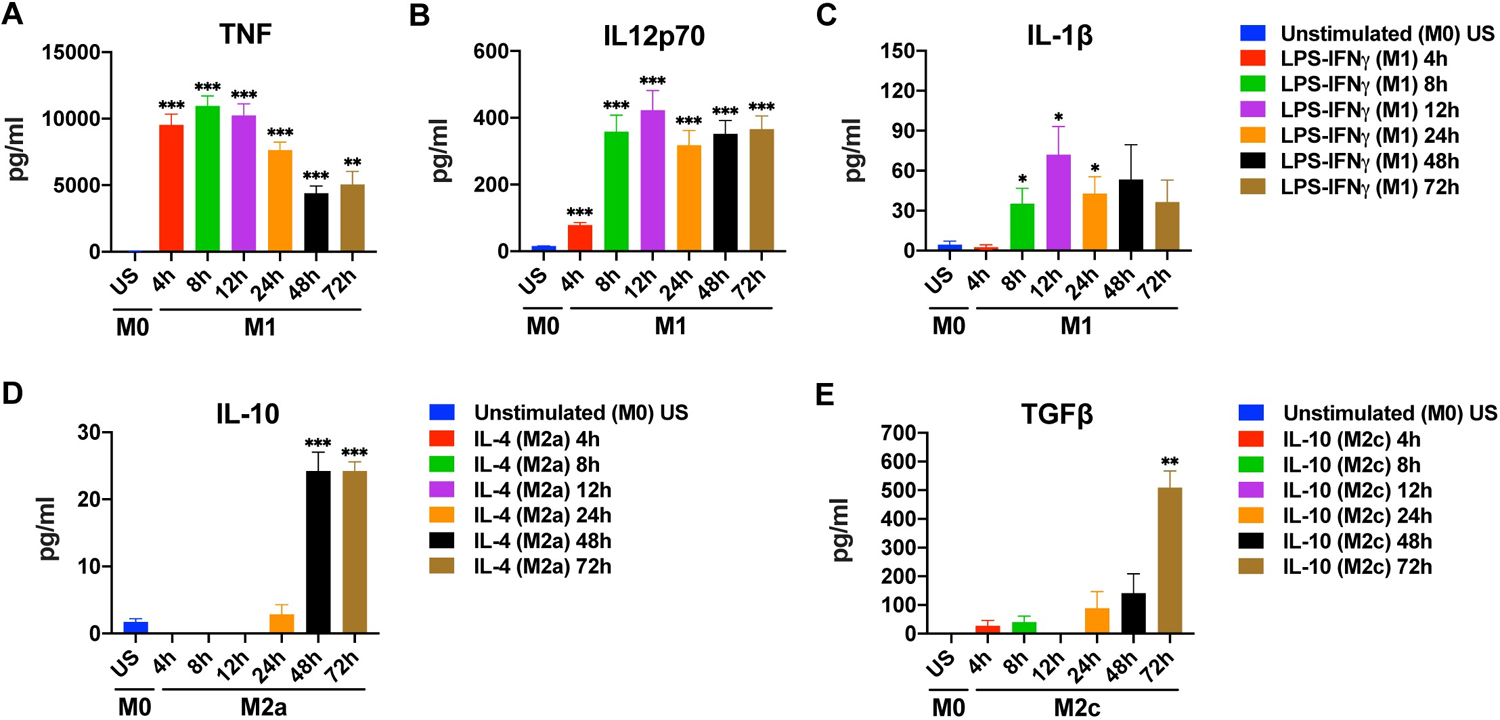
Figure 3. Time-dependent changes in cytokine production by M1, M2a, and M2c macrophages
4. Additional Influences: Cell Type and Species Differences
The study further confirmed that temporal effects are not universally applicable but are modulated by cell type and species. The time course patterns of human primary macrophages (MDMs) differ markedly from those of commonly used cell lines like THP 1—for example, THP 1 shows weaker responses to LPS and opposite IL-10 expression patterns compared to primary macrophages. This implies that "time rules" derived from one cell type cannot be directly extrapolated to other cell types or species.
In summary, macrophage polarization markers do not follow a uniform, stable temporal expression pattern. Even under identical polarization conditions, different molecules exhibit varied trends at the transcriptional, surface-expression, and secretion levels, with clear disparities in their peak time windows. This suggests that discrepancies among studies regarding whether a marker is "upregulated" or its "degree of upregulation" may not stem solely from experimental systems or detection methods, but could be closely tied to the chosen time point of detection. Relying on a single time point may fail to capture the dynamic nature of polarization and introduce uncertainty in data interpretation.
III. Research Implications: New Dimensions for Scientific Inquiry
This study on the "potential impact of time" does not prescribe a universal "optimal detection time" standard, nor does it directly instruct experimental operations. However, it offers valuable conceptual insights for macrophage polarization research, especially when experiments encounter bottlenecks:
1. Interpreting Experimental Variability: An Additional Troubleshooting Dimension
When results diverge between batches or laboratories, beyond routine checks of cell status, reagent concentration, and operational procedures, consider whether "the detection time aligns with the expression rhythm of the marker". The discrepancy might not reflect flawed experimental design, but simply missing the marker’s expression peak. For example, Lab A detecting M1-derived CXCL9 (an early marker) at 4 h obtains high expression, while Lab B testing the same marker at 24 h naturally sees a difference.
2. Troubleshooting False Negatives: Broadening the Perspective
When expected polarization markers are not detected, do not immediately conclude polarization failure. Instead, consider whether "an appropriate detection time was selected." For instance, low CD206 expression in M2a at 24 h may not indicate failed polarization—this marker peaks at 48–72 h. Similarly, low IL-12 expression in M1 at 24 h could reflect its "trough between two peaks."
3. Mechanistic Exploration: Using "Temporal Rhythm" as a New Entry Point
Why do different markers exhibit distinct temporal rhythms? Which signaling pathways regulate these rhythms? For example, does the bimodal pattern of IL-12 relate to transcription-factor binding kinetics? Does the transcription-translation delay of CD206 in M2a involve post-translational modifications or intracellular trafficking? Exploring such questions will deepen our understanding of the dynamic regulatory network underlying macrophage polarization and open new avenues for mechanistic studies.
4. Experimental Design: Incorporating Temporal Flexibility
Although no universal time standard yet exists, future studies could incorporate temporal considerations based on research objectives. For key markers, setting multiple time points to observe dynamic changes—rather than relying on a single snapshot—can prevent time-related bias and provide a more comprehensive view of the polarization process, strengthening research conclusions.
IV. Conclusion
Macrophage polarization is not a "static endpoint" but a "dynamic process" modulated by time. As a potential variable, the influence of time on polarization marker expression offers a fresh perspective for interpreting experimental variability and troubleshooting bottlenecks.
Currently, no unified conclusion exists regarding the effect of time on macrophage polarization, and related research remains exploratory. Nevertheless, this does not diminish its value as an important reference. For researchers, awareness of this potential influence enables more comprehensive consideration of variables, more rational interpretation of results, and may even reveal new entry points for mechanistic studies. As more research unfolds, we may gradually unravel the deeper mechanisms of time-dependent polarization regulation, providing more precise support for macrophage research.
abinScience Products for Macrophage Detection
1. Flow Cytometry Antibodies
2. Recombinant Proteins
3. Other Antibodies
| Target | Reactivity | Product Name | Application | Catalog No. |
|---|---|---|---|---|
| iNOS | Human | Anti-NOS2 Polyclonal Antibody | ELISA, IHC, WB | HW388014 |
| Arg-1 | Human, Mouse, Rat | Anti-ARG1/Arginase-1 Polyclonal Antibody | ELISA, IHC, WB | HY339024 |
| Human, Mouse, Rat, Pig, etc | Anti-ARG1 Polyclonal Antibody | ELISA, IHC, WB | HY339014 | |
| CD11b | Human | Anti-CD11b/ITGAM Polyclonal Antibody | ELISA, IHC, WB | HY474014 |
| Mouse | Anti-Mouse CD11b/ITGAM Polyclonal Antibody | ELISA, IHC, WB | MY474014 | |
| CD86 | Human | Anti-CD86 Polyclonal Antibody | ELISA, IHC, WB | HW776014 |
| Mouse | Anti-Mouse CD86/B7-2 Polyclonal Antibody | ELISA, IHC, WB | MW776014 | |
| CD206 | Human, Mouse, Rat | Anti-CD206/MRC1 Polyclonal Antibody | ELISA, IHC, WB | HB976014 |
| IL-1 | Human, Cercocebus atys, Macaca fascicularis, etc | Anti-IL1B/IL1F2 Polyclonal Antibody | ELISA, IHC, WB | HF943014 |
| IL-6 | Human | Anti-Human IL6 Antibody | ELISA, FCM, WB, IHC, IF | HY328033 |
| Mouse | Anti-Mouse IL6 Monoclonal Antibody | ELISA, IHC, WB | MY328085 | |
| IL-10 | Human | Anti-Human IL10 Antibody | ELISA, WB, IHC, FCM | HB997023 |
| TNF-α | Human, Dog, Cat, Pig, etc | Anti-TNFa/TNF-alpha Polyclonal Antibody | ELISA, IHC, WB | HF879014 |
| Mouse, Rat, Peromyscus leucopus | Anti-TNFa/TNF-alpha Polyclonal Antibody | ELISA, IHC, WB | MF879014 | |
| Danio rerio | Anti-Zebrafish TNFa Polyclonal Antibody | ELISA, IHC, WB | ZA439014 | |
| TGF‐β | Human, Mouse, Dog, Rat, etc | Anti-TGFB1/TGF-beta-1 Polyclonal Antibody | ELISA, IHC, WB | HF977014 |
| Mouse | Anti-Mouse TGFB1/TGF-beta-1 Polyclonal Antibody | ELISA, IHC, WB | MF977014 |
abinScience offers a comprehensive range of antibodies and recombinant proteins for macrophage research, widely applicable in experiments such as ELISA, Western Blot, immunohistochemistry, and flow cytometry. These products support researchers in in-depth studies of macrophage polarization, activation, protein expression, functions, and phenotypes. For more macrophage-related products, visit the abinScience official website: www.abinscience.com.
References
Unuvar Purcu D, Korkmaz A, Gunalp S, Helvaci DG, Erdal Y, Dogan Y, Suner A, Wingender G, Sag D. Effect of stimulation time on the expression of human macrophage polarization markers. PLoS One. 2022 Mar 14;17(3):e0265196. doi: 10.1371/journal.pone.0265196. PMID: 35286356; PMCID: PMC8920204.
+86-027-65523339
중국 우한시 심둔사로 666번지 C동, 우한, 430206





 中文
中文 English
English 한국어
한국어 日本語
日本語 Español
Español Français
Français Русский
Русский