Macroautophagy is one of the cell’s core defenses against invading pathogens. Autophagosomes fuse with lysosomes to form autolysosomes, degrading and clearing intracellular pathogens and damaged components. This fusion step depends on the GTPase RAB7 recruiting the HOPS complex, which in turn drives assembly of the STX17–SNAP29–VAMP8 SNARE complex — a critical checkpoint in the entire pathway.
African swine fever virus (ASFV) remains one of the most severe infectious threats facing the global pig industry, with mortality approaching 100% and no safe, effective vaccine or treatment currently available. ASFV is known to suppress host autophagy, but the underlying mechanism has been unclear. A research team from the College of Veterinary Medicine, Shandong Agricultural University, published a study in Autophagy focusing on the ASFV-encoded I10L protein, systematically mapping out how it interferes with autophagy and exploring its functional significance for viral replication.
I10L: A Key Autophagy-Suppressing Protein Encoded in the ASFV Right Variable Region
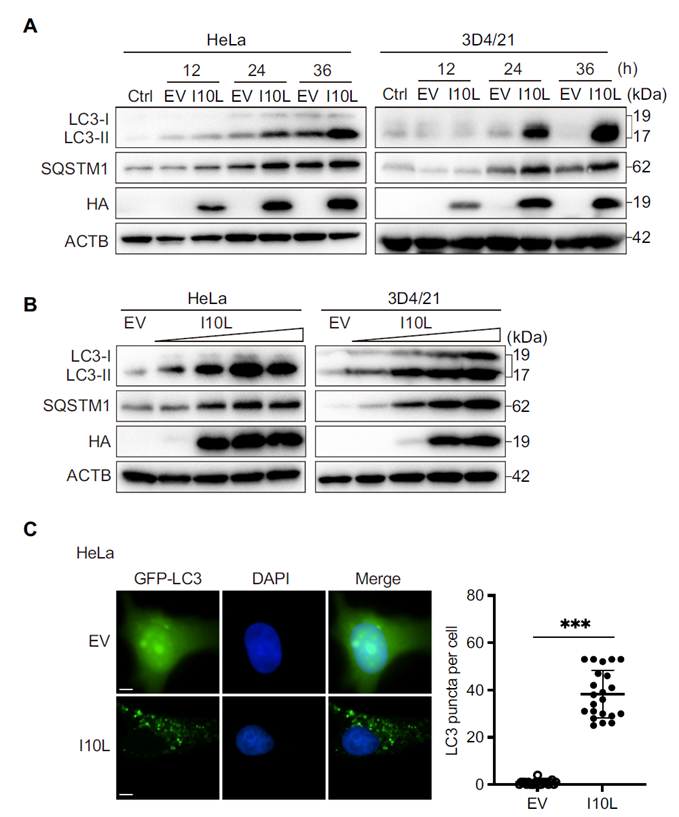
The researchers carried out a systematic screen of ten proteins encoded in the right variable region of the ASFV genome (L11L, I10L, I9R, I8L, and others), overexpressing each in HeLa cells and measuring changes in the autophagy markers LC3-II and SQSTM1/p62. I10L stood out, significantly increasing levels of both LC3-II and SQSTM1/p62 in a time- and dose-dependent manner. GFP-LC3 puncta assays further confirmed that I10L expression markedly increased the number of autophagosomes within cells.
I10L Blocks the Fusion Step of Autophagic Flux, Not Autophagy Initiation
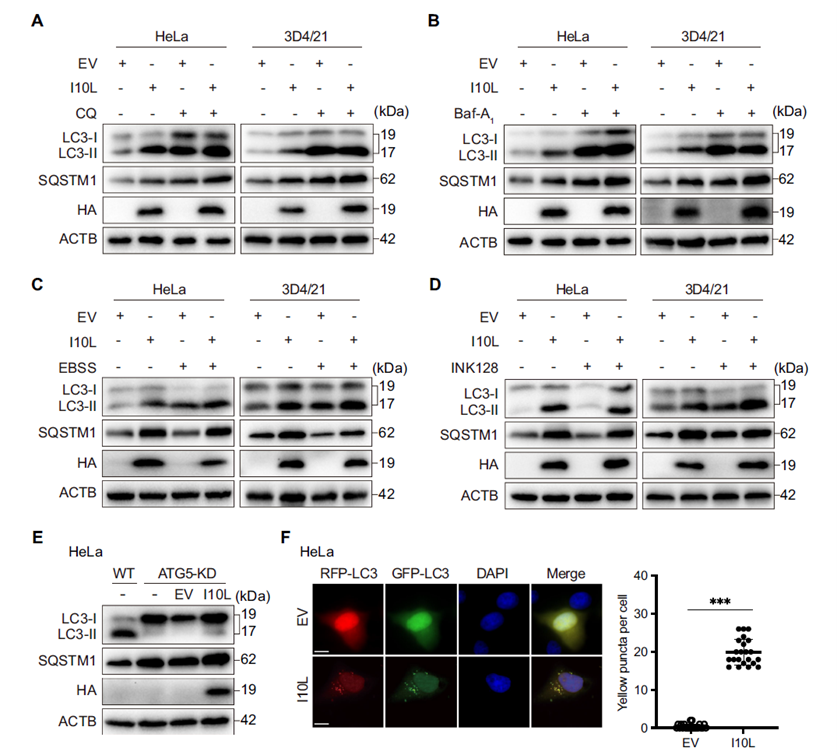
To distinguish between “enhanced autophagy initiation” and “blocked autophagic flux,” the researchers used chloroquine (CQ) and bafilomycin A1 (Baf-A1) to inhibit autophagosome–lysosome fusion, and EBSS starvation or INK128 treatment to activate autophagy initiation. When autophagic flux was already blocked by CQ or Baf-A1, additional I10L expression did not further raise LC3-II or SQSTM1/p62 levels, suggesting I10L acts at the same node as these inhibitors. The same lack of additive effect was observed under ATG5 knockdown. Using the RFP-GFP-LC3 dual fluorescent reporter to track autophagic flux, cells expressing I10L showed a marked accumulation of yellow puncta — representing immature autophagosomes — directly confirming that I10L blocks fusion between autophagosomes and lysosomes.
I10L-Mediated Autophagy Blockade Requires Cooperation Between Its Transmembrane Domain and C-Terminal Region
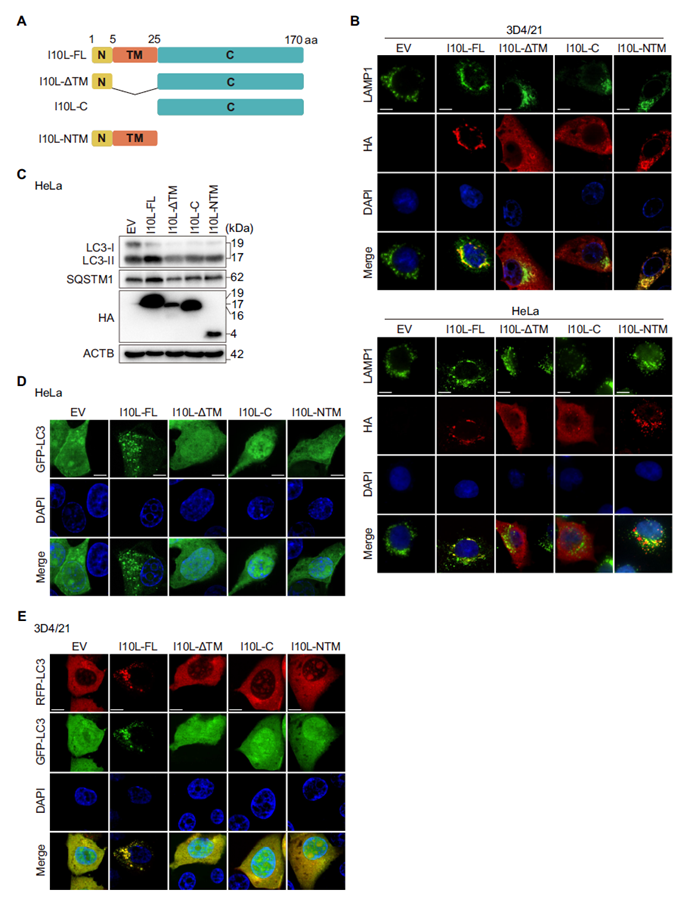
The full-length I10L protein is 170 amino acids long, comprising a short N-terminal segment, a transmembrane domain (TM), and a C-terminal functional region. Immunofluorescence localization showed that both full-length I10L and a truncated construct containing only the TM domain (I10L-NTM) localized to LAMP1-positive lysosomal compartments, whereas the TM-deleted truncation showed no such localization. Functional assays demonstrated that deleting either the TM domain or the C-terminal region alone completely abolished the autophagy-inhibitory effect. The TM domain anchors I10L to the lysosome, while the C-terminus carries out the effector function — both are indispensable.
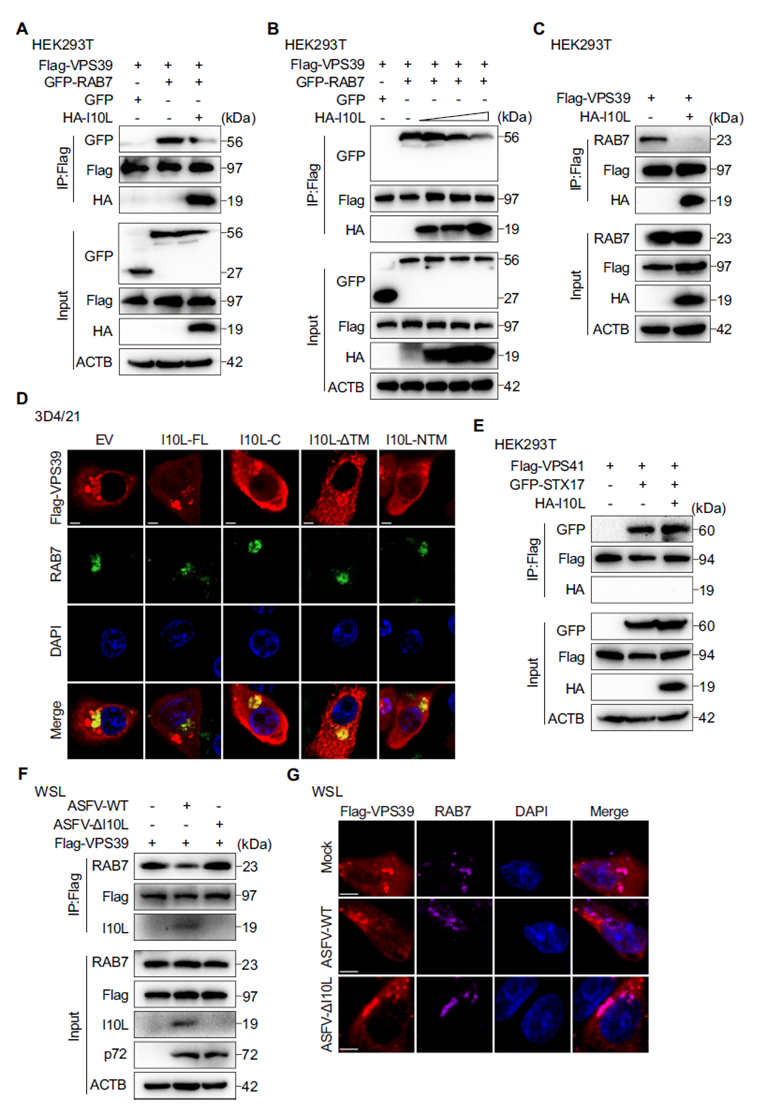
I10L Specifically Binds RAB7, a Marker of Late Endosomes/Lysosomes
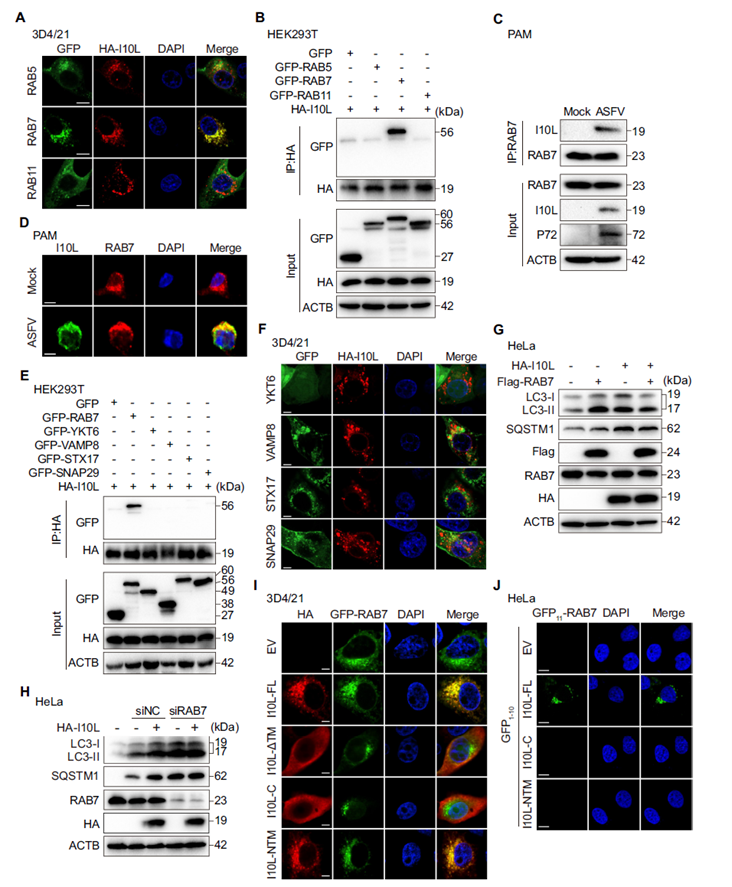
RAB7 is a core regulatory GTPase on late endosomes/lysosomes that serves as an “anchoring bridge” during autophagosome–lysosome fusion. The researchers found that I10L colocalizes extensively with RAB7-marked late endosomes/lysosomes, but not with RAB5 (early endosomes) or RAB11 (recycling endosomes). Co-IP experiments confirmed that I10L binds specifically to RAB7, with no interaction with RAB5, RAB11, or other SNARE proteins. In ASFV-infected primary porcine alveolar macrophages (PAMs), endogenous I10L also interacted and colocalized with RAB7. Overexpressing RAB7 alleviated I10L-induced accumulation of autophagy markers, while RAB7 knockdown prevented I10L from further enhancing the autophagy blockade — demonstrating that I10L exerts its inhibitory effect by hijacking RAB7. Split-fluorescence assays further mapped the interaction interface precisely to amino acids 67–148 in the C-terminus of I10L.
I10L Competitively Occupies RAB7, Blocking Its Binding to the HOPS Subunit VPS39
The HOPS complex binds RAB7 through its VPS39 subunit to drive membrane fusion at late endosomes/lysosomes. Co-IP results showed that I10L expression significantly weakened the interaction between RAB7 and VPS39 in a dose-dependent manner. Immunofluorescence further confirmed that full-length I10L markedly reduced colocalization between VPS39 and endogenous RAB7, an effect not seen with either the TM or C-terminal truncations alone. Notably, I10L did not affect the interaction between the HOPS subunit VPS41 and STX17, indicating that I10L specifically targets the RAB7–VPS39 interface rather than disrupting the entire HOPS complex. Infection with ASFV-ΔI10L restored the RAB7–VPS39 interaction, further supporting this finding.
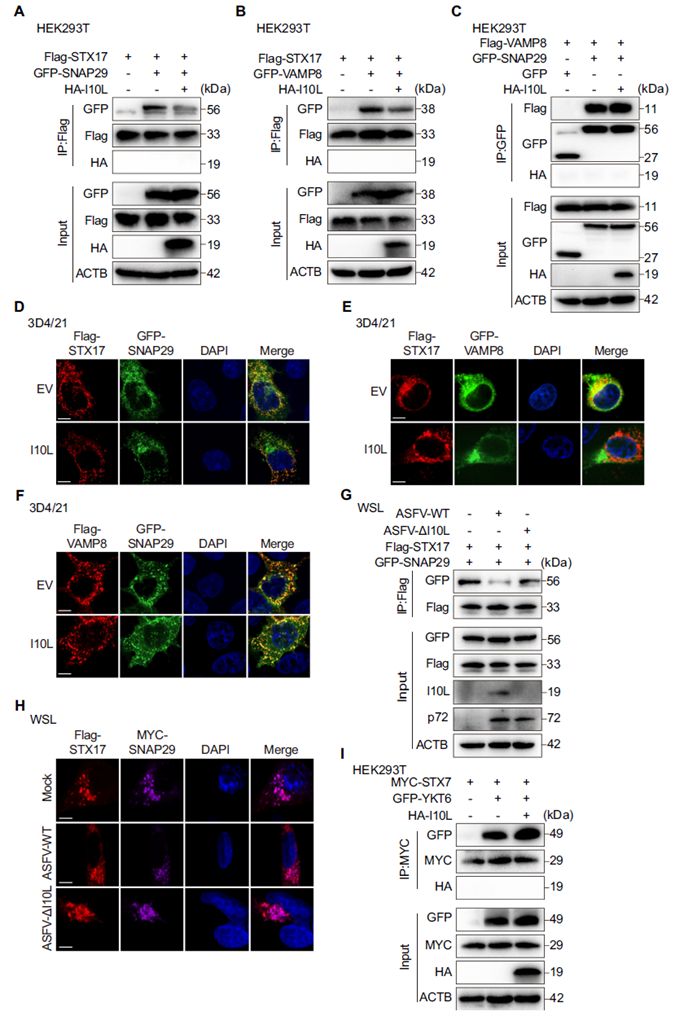
I10L Selectively Disrupts Assembly of the STX17–SNAP29–VAMP8 SNARE Complex
Disruption of the RAB7–HOPS axis ultimately prevents proper assembly of the downstream SNARE complex. Co-IP experiments showed that I10L expression significantly reduced the interactions between STX17 and SNAP29, and between STX17 and VAMP8, while leaving the direct interaction between VAMP8 and SNAP29 unaffected — a pattern consistent with the immunofluorescence colocalization data. In ASFV infection models, the STX17–SNAP29 interaction was markedly reduced in cells infected with ASFV-WT, but this effect was absent with ASFV-ΔI10L. I10L did not interfere with the YKT6–SNAP29–STX7 complex, highlighting its high selectivity for the STX17 pathway.
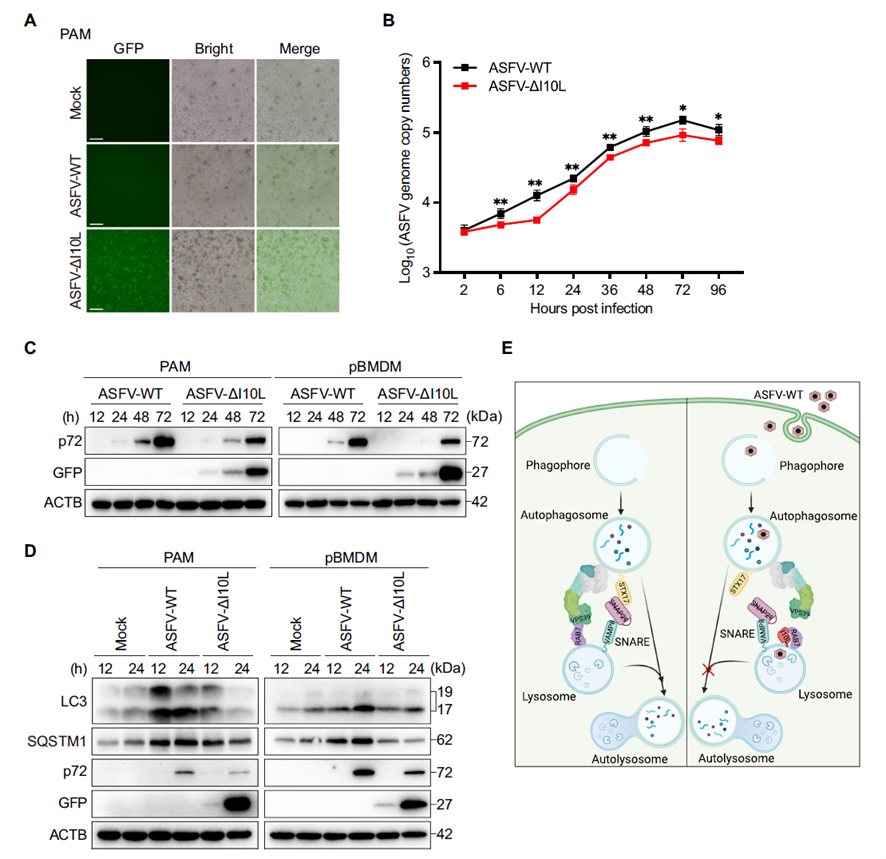
I10L Deletion Significantly Impairs ASFV Replication in Primary Macrophages
When ASFV-ΔI10L and the parental ASFV-WT virus were used to infect PAMs and pBMDMs, qPCR and Western blot both showed that ASFV-ΔI10L replicated significantly less efficiently, with lower viral genome copy numbers and p72 protein levels than the wild-type virus. LC3-II and SQSTM1/p62 levels declined in cells infected with ASFV-ΔI10L, indicating that autophagic flux was partially restored once I10L was removed. Based on these findings, the study proposes an overall mechanistic model: early in infection, ASFV expresses I10L, which competitively binds RAB7, blocks HOPS assembly, and consequently disrupts SNARE complex formation — causing autophagosomes to accumulate without being degraded and thereby promoting ASFV replication.
This study provides a systematic account of how the ASFV-encoded I10L protein — by competitively binding RAB7, blocking HOPS complex assembly, and disrupting SNARE complex formation — inhibits autophagosome–lysosome fusion, allowing the virus to replicate efficiently within an autophagy-blocked cellular environment. These findings identify an important molecular target for understanding ASFV immune evasion and provide a theoretical basis for future antiviral strategies and vaccine development.
Experimental Support from abinScience
abinScience supplied this study with a commercial anti-ASFV I10L antibody – the Anti-ASFV I10L Monoclonal Antibody (1A016) (Catalog No. VK445015). The research team first systematically validated the antibody’s specificity and reactivity, confirming strong performance in both Western blot and immunofluorescence assays. The antibody was then used in endogenous co-IP experiments in ASFV-infected PAMs, successfully detecting the interaction between I10L and endogenous RAB7, and in immunofluorescence to confirm their colocalization in infected cells. These two experiments were essential for validating the I10L–RAB7 interaction under physiologically relevant infection conditions, building on the earlier overexpression-based findings and providing critical support at the level of authentic viral infection.
Need Research Tools for ASFV Studies?
abinScience offers a range of antibodies for ASFV structural proteins, autophagy pathway markers (LC3, p62, RAB7), and SNARE complex components, supporting Western blot, immunofluorescence, co-IP, and viral infection model studies.
Email: info@abinscience.com | Phone: +86-27-65523339

 中文
中文 English
English 한국어
한국어 日本語
日本語 Español
Español Français
Français Русский
Русский