From EV-A71 to CVA6/CVA10: An Overview of Hand, Foot, and Mouth Disease-Associated Enterovirus Research
Release date:
2026-06-18 View count: 115
Ahead of the summer of 2026, hand, foot, and mouth disease (HFMD) has already exhibited an earlier-than-usual surge across the Asia-Pacific region. According to data from the National Institute of Health and Crisis Management of Japan, cumulative confirmed HFMD cases in Japan have reached 11,600 so far this year. During the week of May 25–31, the number of cases admitted to sentinel medical institutions in six prefectures, including Fukuoka, Miyazaki, and Kagoshima, exceeded warning thresholds, prompting local authorities to issue consecutive outbreak alerts significantly earlier than in previous years. The geographic scope of HFMD epidemics extends far beyond the Asia-Pacific region; periodic outbreaks have been recorded in North America and Europe, making it a high-priority infectious disease for global public health surveillance. In China alone, over 15 million cumulative cases were reported between 2013 and 2019, including more than 77,000 severe cases.
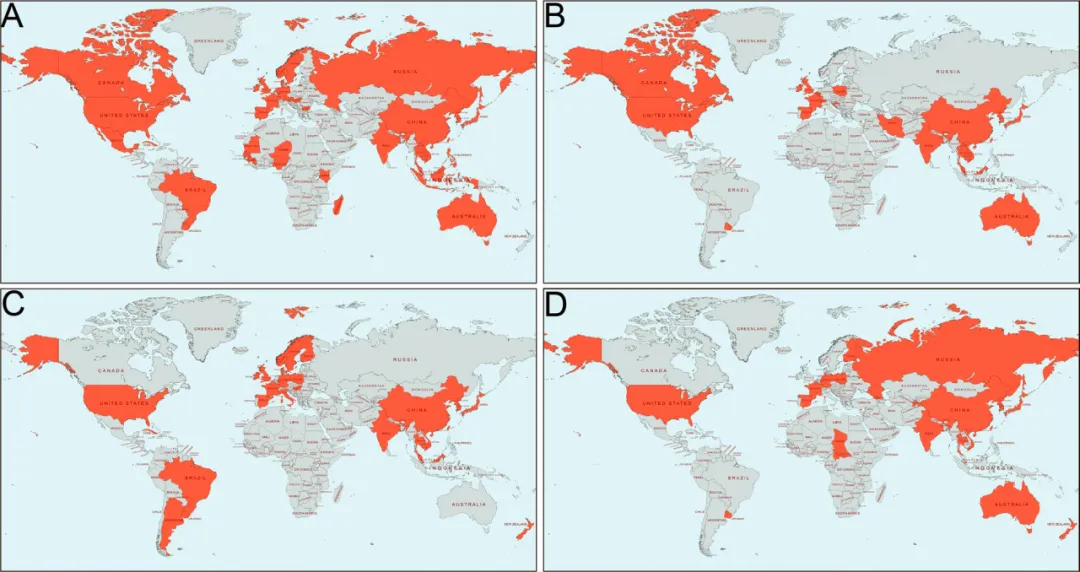
Figure 1. Global distribution of HFMD cases,A EV-A71; B CVA16; C CVA6; D CVA10 (Source: DOI: 10.1186/s12929-023-00908-4)
From a research perspective, the significance of HFMD lies not only in its epidemic scale. The high diversity of its pathogens, the dynamic succession of dominant serotypes, the complexity of spathogenesis of severe disease, and the technical hurdles in multivalent vaccine development have made it an enduring hotspot in virology, immunology, and drug discovery.
Starting with pathogen classification and genetic characteristics, this article systematically reviews recent molecular epidemiology dynamics, insights into pathogenic mechanisms, and cutting-edge advances in vaccines and antiviral therapeutics, aiming to provide a comprehensive reference for researchers in related fields.
1. The Pathogen Landscape: From EV-A71 Dominance to Co-Circulation of Multiple Serotypes
Major Pathogens and Core Characteristics
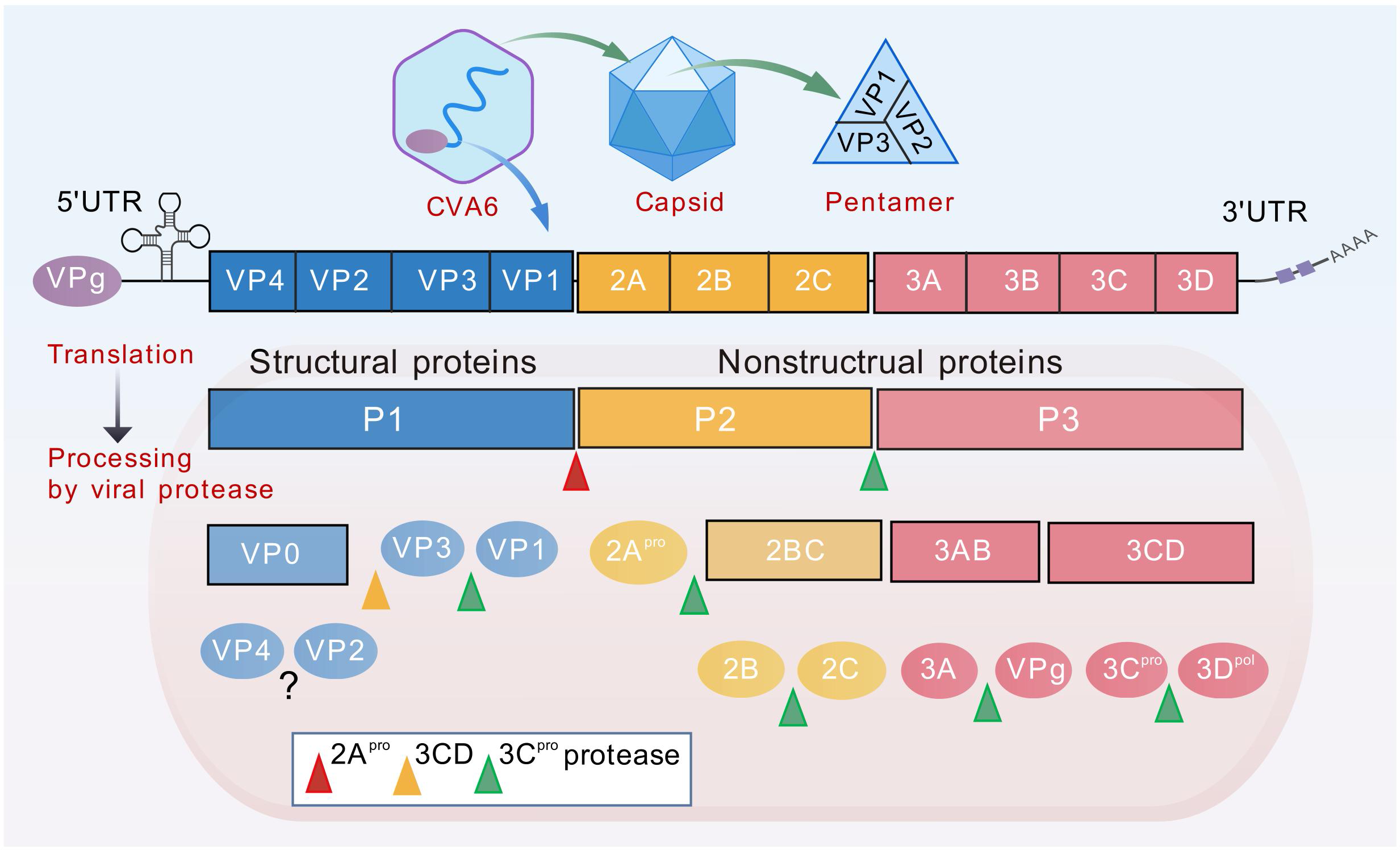
The pathogens responsible for HFMD belong to the genus Enterovirus within the family Picornaviridae. To date, more than 30 serotypes have been confirmed to cause disease, with EV-A71, CVA16, CVA6, and CVA10 serving as the core epidemic strains. These serotypes are all single-stranded, positive-sense RNA (+ssRNA) viruses characterized by non-enveloped, icosahedral spherical particles roughly 20–30 nm in diameter. The full-length genome is approximately 7.4–7.5 kb, flanked by conserved untranslated regions (UTRs) at both ends. It contains a single open reading frame (ORF) that is translated into a polyprotein precursor, which is subsequently cleaved by viral proteases into two main regions: P1 (capsid proteins) and P2/P3 (non-structural proteins).
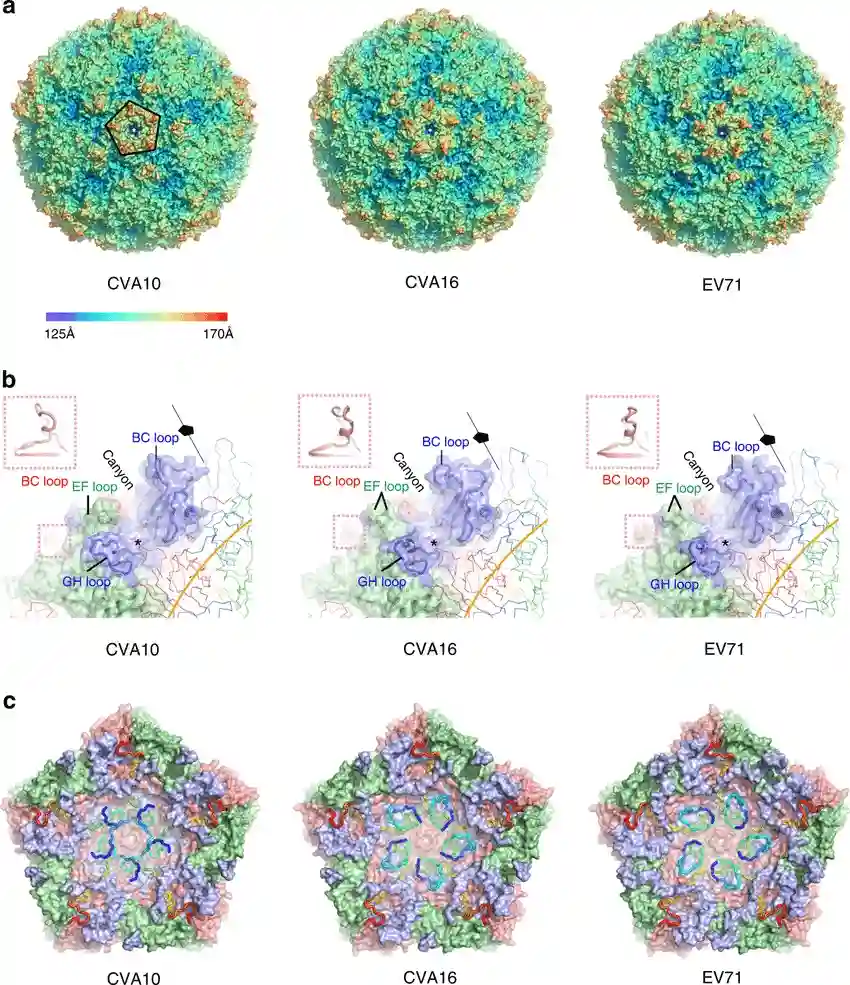
Figure 2. Structural of HFMD pathogens (Source: DOI: 10.1038/s41467-018-07531-0)
These four predominant serotypes display distinct variations in neuroinvasiveness, which serves as a critical determinant of clinical severity:
EV-A71: Exhibits the strongest neurotropism. Infection is often accompanied by persistent high fever, and the virus can invade the central nervous system (CNS), leading to severe complications such as encephalitis, brainstem encephalitis, and neurogenic pulmonary edema. It remains the primary pathogen driving severe and fatal HFMD cases globally and is the most deeply researched HFMD virus in basic science.
CVA16: Historically accounts for the majority of infections historically. Symptoms are relatively mild, and CNS involvement is rare. However, severe and fatal cases attributed to CVA16 have progressively risen across several countries in recent years, indicating that its pathogenic potential should not be underestimated.
CVA6: Has triggered multiple outbreaks worldwide since 2008 and has emerged as the dominant strain in many regions. While its neurotoxicity is lower than that of EV-A71, CVA6 induces more extensive cutaneous lesions. The rash is not restricted to the classic sites (hands, feet, mouth) but can spread across the trunk and extremities. Desquamation and nail shedding (onychomadesis) are frequently observed during the convalescent phase, presenting new challenges for differential clinical diagnosis. Additionally, CVA6 is highly prone to genetic recombination, leading to continuously expanding genotypic diversity and driving the current evolution of the pathogen spectrum.
CVA10: Frequently co-circulates with CVA6. Its detection rate in clinical samples from severe patients has risen notably in certain regions recently, drawing increased research attention.
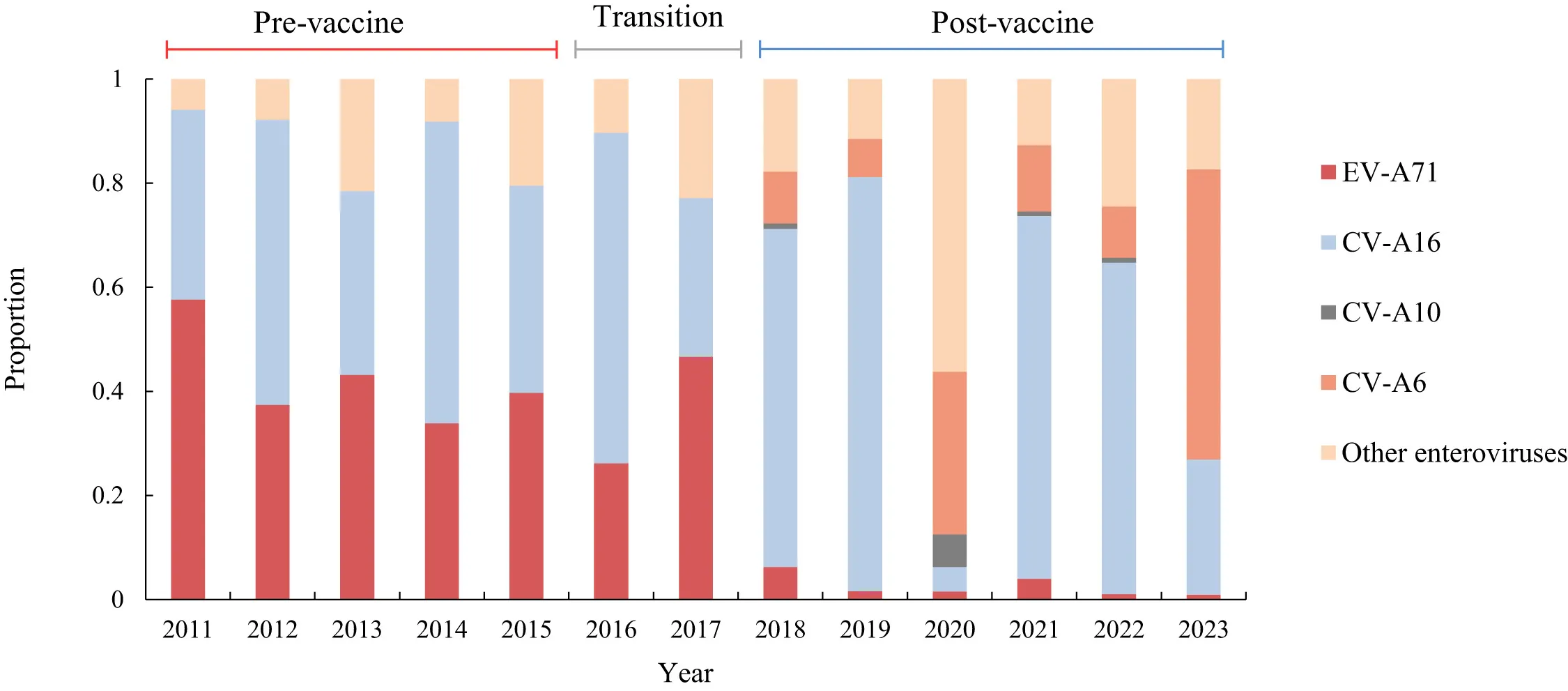
Figure 3. Shifts in viral pathogen distribution (Source: DOI: 10.1016/j.lanwpc.2025.101603)
2. Structure and Function of Viral Proteins: A Panoramic View of Research Targets
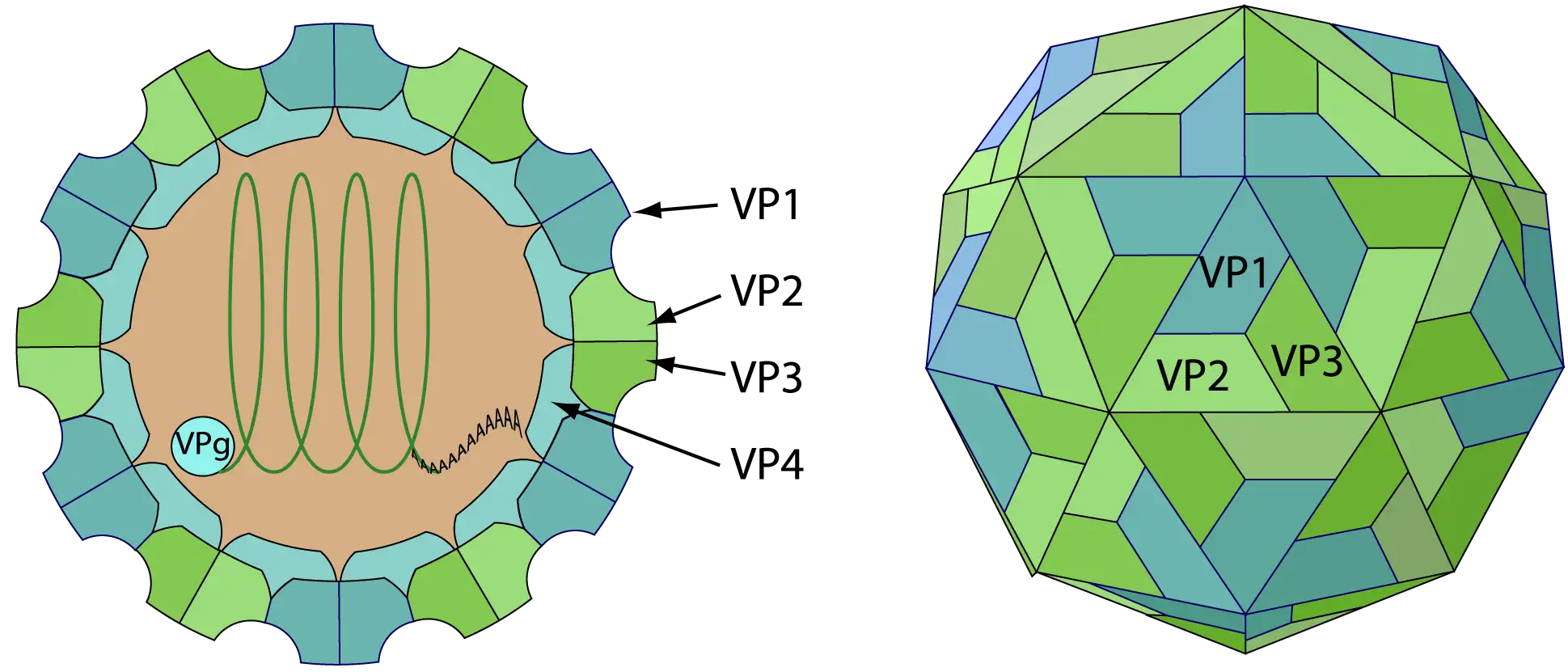
The enterovirus capsid is assembled from 60 copies each of four structural proteins: VP1, VP2, VP3, and VP4. Among them, VP1, VP2, and VP3 are exposed on the particle surface, forming the molecular interface that interacts with the external environment, including host receptors and neutralizing antibodies. Conversely, VP4 resides on the inner surface of the capsid and plays a pivotal role in viral uncoating.
Structural Protein
Location
Primary Function
Research & Drug Development Significance
VP1
Capsid surface
Recognizes and binds to the host receptor SCARB2; carries primary neutralizing epitopes.
Primary target for vaccine immunogen design, neutralizing antibody screening, and serotype identification.
VP2
Capsid surface
Maintains capsid structural stability; participates in the formation of secondary neutralizing epitopes.
Broad-spectrum neutralizing antibody discovery; cross-protection mechanisms of multivalent vaccines.
VP3
Capsid surface
Maintains capsid stability; participates in viral particle assembly.
Mechanisms of viral particle assembly; target for capsid inhibitors.
VP4
Inner capsid
Mediates viral RNA release from endosomes during uncoating.
Viral entry pathway analysis; development of uncoating inhibitors.
Among these, VP1 is the most widely utilized target protein in HFMD research. Amplification and sequencing of the ~900 bp region within VP1 serve as the gold-standard method for enterovirus serotyping and genotyping. In vaccine development, recombinant VP1 proteins or virus-like particles (VLPs) featuring VP1 as a core component are primary immunogen candidates for multivalent subunit vaccines. In immunological studies, the VP1 protein is also a vital tool for screening monoclonal antibodies with neutralizing activity and evaluating vaccine-induced humoral immunity.
Additionally, VP2 harbors certain antigenic epitopes that are relatively conserved between EV-A71 and CVA16. This provides a strategic entry point for discovering broad-spectrum neutralizing antibodies; targeting these conserved epitopes could inform the design of multivalent vaccines and passive immunization strategies.
Figure 5. Diagram of CVA6 genome structure (Source: DOI: 10.3389/fimmu.2025.1603028)
The non-structural proteins encoded by the P2 and P3 regions are not only responsible for viral RNA replication and polyprotein processing/maturation but also disrupt normal host cell functions and immune responses through multiple mechanisms. They form the molecular foundation for efficient viral replication and immune evasion.
Non-Structural Protein
Primary Function
Research & Drug Development Value
2Apro (Protease)
Cleaves the polyprotein precursor; cleaves host translation initiation factor eIF4G to shut down host protein synthesis; cleaves MAVS to block the type I interferon (IFN-I) pathway.
Immune evasion mechanism studies; target for protease inhibitor development.
2B
Enhances cell membrane (ER/Golgi) permeability to promote viral particle release; participates in modulating cellular calcium homeostasis.
Viral egress and release mechanism studies.
2C
Displays NTPase/helicase activity; involved in membranous vesicle formation and assembly of replication organelles (replication complexes).
Viral replication organelle research; broad-spectrum antiviral target (e.g., target for certain inhibitors like chloroquine).
3A
Inhibits intracellular vesicle trafficking; participates in anchoring the replication complex to membranes.
Virus-host interaction studies.
3B(VPg)
Covalently links to the 5' end of genomic RNA, serving as a primer for RNA synthesis to initiate genome replication.
Viral RNA replication mechanism studies.
3Cpro (Protease)
Principal protease executing polyprotein cleavage; cleaves TRIF to disrupt TLR signaling pathways; cleaves host transcription factors to suppress gene expression.
The most mature broad-spectrum drug target against HFMD; multiple inhibitors are under active investigation.
3Dpol (Polymerase)
RNA-dependent RNA polymerase (RdRp) that executes viral genome replication.
Target for nucleoside analog antivirals; screening of polymerase inhibitors.
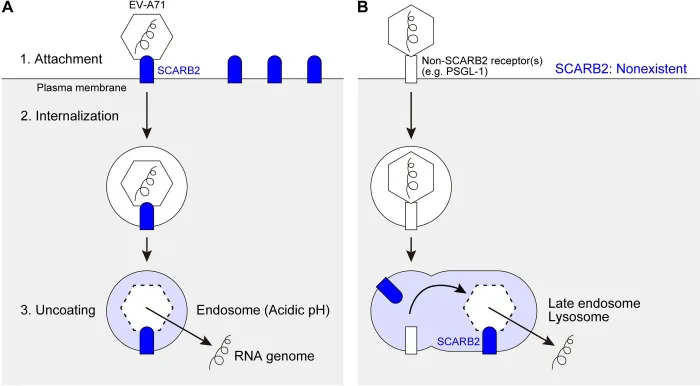
Elucidating viral receptors is a central theme in enterovirus pathogenesis research. For EV-A71, several receptors and co-receptors have been identified:
hSCARB2 (Human Scavenger Receptor Class B Member 2): Serves as the primary functional receptor for EV-A71, capable of recognizing all EV-A71 strains and mediating viral uncoating upon entry. Interestingly, while its cell-surface expression is relatively low, its primary functionality manifests within the endosomal/lysosomal lumen, acting as an intracellular receptor that triggers the release of viral RNA.
PSGL-1 (P-Selectin Glycoprotein Ligand-1): Identified as the second receptor for EV-A71, though its binding is highly strain-specific. Currently, only about 20% of EV-A71 isolates bind to PSGL-1. Notably, PSGL-1 binding capacity correlates with viral virulence—strains capable of binding PSGL-1 (typically possessing A/G/Q at position 145 of the VP1 protein) are detected with significantly higher frequency in severe clinical cases.
Co-receptors: Molecules such as heparan sulfate proteoglycans (HSPG), annexin A2 (Anx2), vimentin, and fibronectin have also been identified as co-receptors. They primarily facilitate viral attachment or adsorption but cannot independently mediate viral entry in the absence of SCARB2 or PSGL-1.
For CVA6 and CVA10, recent studies have confirmed that SCARB2 also participates in their cellular entry process. This suggests that SCARB2 may function as a broad-spectrum receptor across Enterovirus A species, offering a theoretical foundation for developing host-receptor-targeted pan-enterovirus antiviral strategies.
3.2 Immune Evasion and Innate Immune Antagonism
During infection, enteroviruses actively counteract host innate immune responses via their encoded protease machinery. The proteases 2Apro and 3Cpro serve as the primary tools for immune antagonism. Both can proteolytically cleave MAVS (a critical adaptor protein in the RIG-I/MDA5 signaling pathway) and TRIF (an adaptor protein in the TLR signaling pathway), thereby blocking the production of type I interferons (IFN-I). Because IFN-I is the core effector molecule of antiviral innate immunity, this active "shielding" mechanism significantly extends the replication window of the virus within the host.
In the pathogenesis of severe cases, dysregulation of the host immune response—rather than viral load alone—is recognized as the critical driver. Studies indicate that patients with severe EV-A71 infections experience systemic cytokine storms and a disrupted Th17/Treg balance. Markedly elevated levels of pathogenic cytokines, such as IL-17A and IFN-γ, correlate closely with the development of neurogenic pulmonary edema and brainstem encephalitis.
4. Antiviral Drug Development: Targets and Strategies
To date, no specific antiviral therapeutics have been approved for marketing against HFMD-associated enteroviruses; clinical management remains heavily reliant on symptomatic and supportive care. This gap represents an urgent clinical need, while simultaneously providing a vast landscape for basic and translational research.
4.1 The 3C Protease: The Most Consensus-Backed Antiviral Target
The 3C protease (3Cpro) exerts dual roles in viral replication: it executes the cleavage of the viral polyprotein precursor (essential for structural and non-structural protein maturation), and it actively suppresses innate immunity by cleaving host signaling proteins. Because 3Cpro is highly conserved across various enterovirus serotypes and shares virtually no homology with human cellular proteins, it stands out as an ideal target for pan-enterovirus antivirals.
4.2 The 3D Polymerase and Capsid Proteins: Supplementary Targets
The 3D polymerase (3Dpol), an RNA-dependent RNA polymerase (RdRp), is the catalytic engine of viral genome replication and serves as a major target for nucleoside analog antivirals.
Additionally, the VP1 capsid protein harbors a hydrophobic pocket (analogous to the binding site of pleconaril in rhinoviruses). The binding of small-molecule ligands to this pocket locks the conformation of the viral particle and prevents uncoating, thereby blocking viral entry. Recent virtual screening campaigns leveraging the DrugBank database alongside structural data for VP1-4, 3Cpro, and 3Dpol have identified several promising candidate compounds, some of which have demonstrated preliminary efficacy in in vitro cell cultures and in vivo murine models.
Summary and Outlook
HFMD represents a quintessential "known but unconquered" landscape in infectious disease research. While robust knowledge has accumulated regarding pathogen classification, genetic evolution, vaccine development, and drug targets, significant gaps remain across several critical dimensions:
1) Development and Optimization of Multivalent Vaccines: Overcoming immune interference among diverse serotypic components, optimizing formulation ratios and adjuvant systems, and establishing a clear dose-response relationship between neutralizing antibody titers and protective efficacy stand as the most pressing and challenging scientific bottlenecks.
2) Translational Advancement of Broad-Spectrum Antiviral Strategies: Although 3Cpro inhibitors and neutralizing antibodies have exhibited promising pan-enterovirus activity, transitioning from in vitro potency to in vivo efficacy and safety demands systematic evaluation in pharmacokinetics/pharmacodynamics (PK/PD), toxicology, and robust animal models.
3) Real-Time Evolutionary Surveillance and Early Warning: With the anticipated rollout of multivalent vaccines, shifts in the pathogen spectrum—similar to those observed following monovalent EV-A71 vaccination—are highly probable. Establishing a real-time, whole-genome sequencing surveillance infrastructure covering major serotypes to capture antigenic drift and novel recombinants promptly is essential for future precision public health interventions.
4) In-Depth Mechanistic Research into CVA6 and CVA10 Pathogenesis: Compared to the wealth of data on EV-A71, research into the receptor-binding dynamics, immune evasion tactics, and severe disease pathways of CVA6 and CVA10 remains in its infancy, marking these as high-value, emerging frontiers for investigation.
abinScience HFMD-Related Research Tools
abinScience has developed recombinant protein and antibody products targeting both structural and non-structural viral proteins for major circulating serotypes of HFMD-associated enteroviruses (EV-A71, CVA16, CVA6, etc.).These products support a wide range of applications, from viral structural analysis, host immune response assessment, neutralizing antibody screening to serotype identification, serving as core reagents to advance basic virology research and translational development for HFMD.
[1]. Zhu, P., Ji, W., Li, D., Li, Z., Chen, Y., Dai, B., Han, S., Chen, S., Jin, Y., & Duan, G. (2023). Current status of hand-foot-and-mouth disease. Journal of biomedical science, 30(1), 15. https://doi.org/10.1186/s12929-023-00908-4
[2]. Ge M, et al. Pre-existing antibodies and age shape the immune response following EV-A71 vaccine: a prospective serological study among the Chinese pediatric population. medRxiv, 2025. https://doi.org/10.1101/2025.09.17.25335968
[3]. Li Y, Ruan Y, Zhai X, et al. Frontiers and hotspots in hand, foot, and mouth disease research during 2006 to 2023: A bibliometric and visual analysis. Medicine, 2024, 103(24): e38550. https://doi.org/10.1097/MD.0000000000038550
[4]. Hu K, Onintsoa Diarimalala R, Yao C, et al. EV-A71 Mechanism of Entry: Receptors/Co-Receptors, Related Pathways and Inhibitors. Viruses, 2023, 15(3): 785. https://doi.org/10.3390/v15030785
[5]. Liu, F., Yi, Y., Song, Y., Zhang, X., Xie, T., Liu, Y., Chen, Y., Huang, S., Zhang, J., Zhang, Y., Chang, Z., & Cui, F. (2025). Epidemiology of hand, foot, and mouth disease outbreaks before and during availability of EV-A71 vaccine in China's mainland: analysis of outbreak surveillance data from 2011 to 2023. The Lancet regional health. Western Pacific, 59, 101603. https://doi.org/10.1016/j.lanwpc.2025.101603
[6]. Nishimura Y, et al. Cellular receptors for enterovirus A71. PLoS Pathogens, 2024, 20(2): e1012022. https://doi.org/10.1371/journal.ppat.1012022
[7]. Tay MYF, et al. The host immune response to enterovirus A71 (EV-A71): from viral immune evasion to immunopathology and prognostic biomarkers of severe disease. Viruses, 2024.
[8]. Lv J, et al. Reversible covalent inhibitors suppress enterovirus 71 infection by targeting the 3C protease. Antiviral Research, 2021, 190: 105075. https://doi.org/10.1016/j.antiviral.2021.105075
[9]. Xu X, et al. A clinical SARS-CoV-2 Mpro inhibitor blocks replication of multiple enteroviruses and confers oral in vivo protection in animal models. PLoS Pathogens, 2026, 22: e1014051. https://doi.org/10.1371/journal.ppat.1014051
[10]. Tan CW, et al. Tanomastat exerts multi-targeted inhibitory effects on viral capsid dissociation and RNA replication in human enteroviruses. eBioMedicine, 2024. https://doi.org/10.1016/j.ebiom.2024.105183
[11]. Khamis MM, et al. Computational screening of newly designed compounds against Coxsackievirus A16 and Enterovirus A71. PMC, 2022.

 中文
中文 English
English 한국어
한국어 日本語
日本語 Español
Español Français
Français Русский
Русский